Respiratory system
The lungs and respiratory system allow us to breathe. They bring oxygen into our bodies (called inspiration, or inhalation) and send carbon dioxide out (called expiration, or exhalation).
This exchange of oxygen and carbon dioxide is called respiration.
Parts of Respiratory system
The respiratory system includes the
nose,
mouth,
throat,
voice box,
windpipe, and
lungs.
Air enters the respiratory system through the nose or the mouth. If it goes in the nostrils (also called nares), the air is warmed and humidified. Tiny hairs called cilia protect the nasal passageways and other parts of the respiratory tract, filtering out dust and other particles that enter the nose through the breathed air.
The two openings of the airway (the nasal cavity and the mouth) meet at the pharynx, or throat, at the back of the nose and mouth. The pharynx is part of the digestive system as well as the respiratory system because it carries both food and air.
At the bottom of the pharynx, this pathway divides in two, one for food — the esophagus, which leads to the stomach — and the other for air. The epiglottis, a small flap of tissue, covers the air-only passage when we swallow, keeping food and liquid from going into the lungs.
The larynx, or voice box, is the top part of the air-only pipe. This short tube contains a pair of vocal cords, which vibrate to make sounds.
The trachea, or windpipe, is the continuation of the airway below the larynx. The walls of the trachea are strengthened by stiff rings of cartilage to keep it open. The trachea is also lined with cilia, which sweep fluids and foreign particles out of the airway so that they stay out of the lungs.
At its bottom end, the trachea divides into left and right air tubes called bronchi, which connect to the lungs. Within the lungs, the bronchi branch into smaller bronchi and even smaller tubes called bronchioles. Bronchioles end in tiny air sacs called alveoli, where the exchange of oxygen and carbon dioxide actually takes place. Each person has hundreds of millions of alveoli in their lungs. This network of alveoli, bronchioles, and bronchi is known as the bronchial tree.
The lungs also contain elastic tissues that allow them to inflate and deflate without losing shape. They're covered by a thin lining called the pleura.
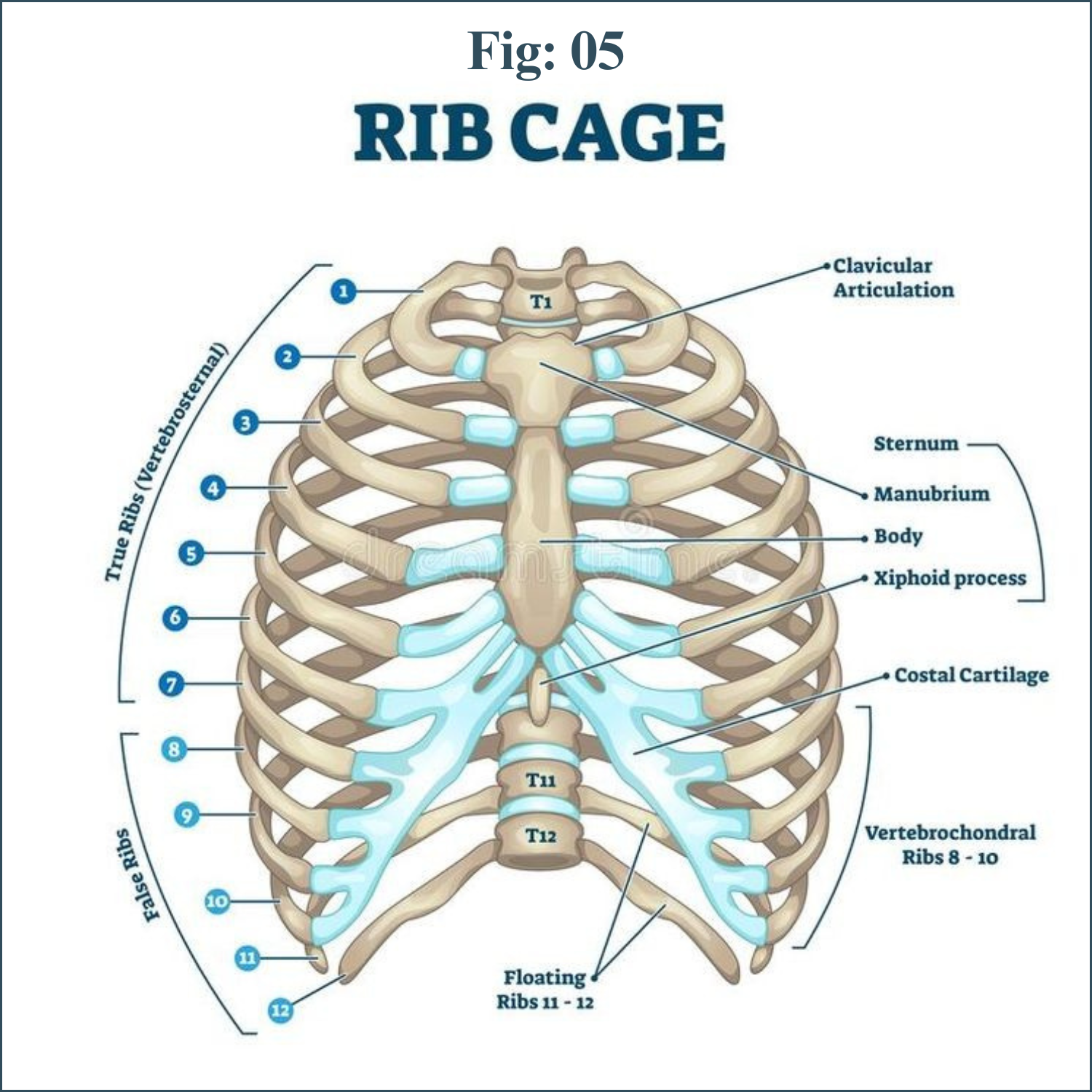
The chest cavity, or thorax, is the airtight box that houses the bronchial tree, lungs, heart, and other structures. The top and sides of the thorax are formed by the ribs and attached muscles, and the bottom is formed by a large muscle called the diaphragm. The chest walls form a protective cage around the lungs and other contents of the chest cavity.

The Gas-exchange Region
The gas-exchange region comprises three compartments: air, blood, and tissue. Whereas air and blood are continuously replenished, the function of the tissue compartment is twofold: it provides the stable supporting framework for the air and blood compartments, and it allows them to come into close contact with each other (thereby facilitating gas exchange) while keeping them strictly confined.
The respiratory gases diffuse from air to blood, and vice versa, through the 140 square metres of internal surface area of the tissue compartment.
The gas-exchange tissue proper is called the pulmonary parenchyma, while the supplying structures, conductive airways, lymphatics, and non-capillary blood vessels belong to the non-parenchyma.
The gas-exchange region begins with the alveoli of the first generation of respiratory bronchioles. Distally, the frequency of alveolar outpocketings increases rapidly, until after two to four generations of respiratory bronchioles, the whole wall is formed by alveoli.
The airways are then called alveolar ducts and, in the last generation, alveolar sacs.
Alveoli
On average, an adult human lung has about 480 million alveoli. They are polyhedral structures, with a diameter of about 250 to 300 μm (1 μm = 0.000039 inch), and open on one side, where they connect to the airway.
The alveolar wall, called the interalveolar septum, is common to two adjacent alveoli. It contains a dense network of capillaries, the smallest of the blood vessels, and a skeleton of connective tissue fibers.
The fiber system is interwoven with the capillaries and particularly reinforced at the alveolar entrance rings. The capillaries are lined by flat endothelial cells with thin cytoplasmic extensions.
The interalveolar septum is covered on both sides by the alveolar epithelial cells. A thin, squamous cell type, the type I pneumocyte, covers between 92 and 95 percent of the gas-exchange surface; a second, more cuboidal cell type, the type II pneumocyte, covers the remaining surface.
The type I cells form, together with the endothelial cells, the thin air–blood barrier for gas exchange; the type II cells are secretory cells.
Type II pneumocytes produce a surface-tension-reducing material, the pulmonary surfactant, which spreads on the alveolar surface and prevents the tiny alveolar spaces from collapsing.
Before it is released into the airspaces, pulmonary surfactant is stored in the type II cells in the form of lamellar bodies.
These granules are the conspicuous ultrastructural features of this cell type. On top of the epithelium, alveolar macrophages creep around within the surfactant fluid.
They are large cells, and their cell bodies abound in granules of various content, partly foreign material that may have reached the alveoli, or cell debris originating from cell damage or normal cell death.
Ultimately, the alveolar macrophages are derived from the bone marrow, and their task is to keep the air–blood barrier clean and unobstructed.
The tissue space between the endothelium of the capillaries and the epithelial lining is occupied by the interstitium. It contains connective tissue and interstitial fluid.
The connective tissue comprises a system of fibers, amorphous ground substance, and cells (mainly fibroblasts), which seem to be endowed with contractile properties. The fibroblasts are thought to control capillary blood flow or, alternatively, to prevent the accumulation of extracellular fluid in the interalveolar septa.
If for some reason the delicate fluid balance of the pulmonary tissues is impaired, an excess of fluid accumulates in the lung tissue and within the airspaces.
This pathological condition is called pulmonary edema. As a consequence, the respiratory gasses must diffuse across longer distances, and proper functioning of the lung is severely jeopardized.


Control of Breathing
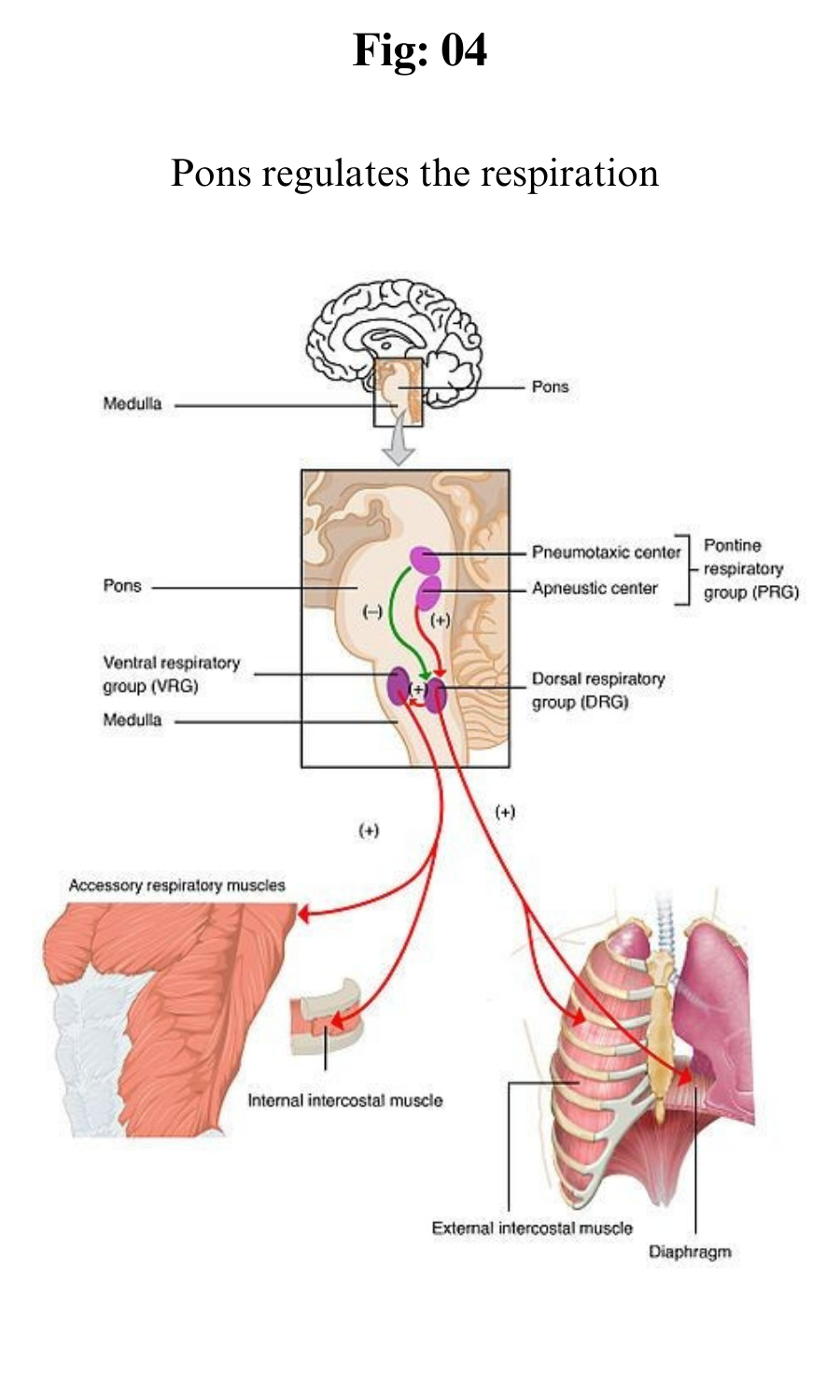
Breathing is an automatic and rhythmic act produced by networks of neurons in the hindbrain (the pons and medulla). The neural networks direct muscles that form the walls of the thorax and abdomen and produce pressure gradients that move air into and out of the lungs.
The respiratory rhythm and the length of each phase of respiration are set by reciprocal stimulatory and inhibitory interconnection of these brain-stem neurons.
An important characteristic of the human respiratory system is its ability to adjust breathing patterns to changes in both the internal milieu and the external environment.
Ventilation increases and decreases in proportion to swings in carbon dioxide production and oxygen consumption caused by changes in metabolic rate.
The respiratory system is also able to compensate for disturbances that affect the mechanics of breathing, such as the airway narrowing that occurs in an asthmatic attack.
Breathing also undergoes appropriate adjustments when the mechanical advantage of the respiratory muscles is altered by postural changes or by movement.
This flexibility in breathing patterns in large part arises from sensors distributed throughout the body that send signals to the respiratory neuronal networks in the brain.
Chemoreceptors detect changes in blood oxygen levels and change the acidity of the blood and brain.
Mechanoreceptors monitor the expansion of the lung, the size of the airway, the force of respiratory muscle contraction, and the extent of muscle shortening.
Although the diaphragm is the major muscle of breathing, its respiratory action is assisted and augmented by a complex assembly of other muscle groups.
Intercostal muscles inserting on the ribs, the abdominal muscles, and muscles such as the scalene and sternocleidomastoid that attach both to the ribs and to the cervical spine at the base of the skull also play an important role in the exchange of air between the atmosphere and the lungs.
In addition, laryngeal muscles and muscles in the oral and nasal pharynx adjust the resistance of movement of gasses through the upper airways during both inspiration and expiration.
Although the use of these different muscle groups adds considerably to the flexibility of the breathing act, they also complicate the regulation of breathing. These same muscles are used to perform a number of other functions, such as speaking, chewing and swallowing, and maintaining posture.
Perhaps because the “respiratory” muscles are employed in performing nonrespiratory functions, breathing can be influenced by higher brain centers and even controlled voluntarily to a substantial degree.
An outstanding example of voluntary control is the ability to suspend breathing by holding one’s breath.
Input into the respiratory control system from higher brain centers may help optimize breathing so that not only are metabolic demands satisfied by breathing but ventilation also is accomplished with minimal use of energy.
Central Organization of Respiratory Neurons
The respiratory rhythm is generated within the pons and medulla oblongata. Three main aggregations of neurons are involved: a group consisting mainly of inspiratory neurons in the dorsomedial medulla, a group made up of inspiratory and expiratory neurons in the ventrolateral medulla, and a group in the rostral pons consisting mostly of neurons that discharge in both inspiration and expiration.
It is thought that the respiratory cycle of inspiration and expiration is generated by synaptic interactions within these groups of neurons.
The inspiratory and expiratory medullary neurons are connected to projections from higher brain centres and from chemoreceptors and mechanoreceptors; in turn they drive cranial motor neurons, which govern the activity of muscles in the upper airways and the activity of spinal motor neurons, which supply the diaphragm and other thoracic and abdominal muscles.
The inspiratory and expiratory medullary neurons also receive input from nerve cells responsible for cardiovascular and temperature regulation, allowing the activity of these physiological systems to be coordinated with respiration.
Neurally, inspiration is characterized by an augmenting discharge of medullary neurons that terminates abruptly. After a gap of a few milliseconds, inspiratory activity is restarted, but at a much lower level, and gradually declines until the onset of expiratory neuron activity.
Then the cycle begins again. The full development of this pattern depends on the interaction of several types of respiratory neurons: inspiratory, early inspiratory, off-switch, post-inspiratory, and expiratory.
Early inspiratory neurons trigger the augmenting discharge of inspiratory neurons. This increase in activity, which produces lung expansion, is caused by self-excitation of the inspiratory neurons and perhaps by the activity of an as yet undiscovered upstream pattern generator.
Off-switch neurons in the medulla terminate inspiration, but pontine neurons and input from stretch receptors in the lung help control the length of inspiration. When the vagus nerves are sectioned or pontine centers are destroyed, breathing is characterized by prolonged inspiratory activity that may last for several minutes. This type of breathing, which occasionally occurs in persons with diseases of the brain stem, is called apneustic breathing.
Post-inspiratory neurons are responsible for the declining discharge of the inspiratory muscles that occurs at the beginning of expiration. Mechanically, this discharge aids in slowing expiratory flow rates and probably assists the efficiency of gas exchange.
It is thought by some that these post-inspiratory neurons have inhibitory effects on both inspiratory and expiratory neurons and therefore play a significant role in determining the length of the respiratory cycle and the different phases of respiration.
As the activity of the post-inspiratory neurons subsides, expiratory neurons discharge and inspiratory neurons are strongly inhibited. There may be no peripheral manifestation of expiratory neuron discharge except for the absence of inspiratory muscle activity, although in upright humans the lower expiratory intercostal muscles and the abdominal muscles may be active even during quiet breathing.
Moreover, as the demand to breathe increases (for example, with exercise), more expiratory intercostal and abdominal muscles contract. As expiration proceeds, the inhibition of the inspiratory muscles gradually diminishes and inspiratory neurons resume their activity.
Chemoreceptors
One way in which breathing is controlled is through feedback by chemoreceptors.
There are two kinds of respiratory chemoreceptors: arterial chemoreceptors, which monitor and respond to changes in the partial pressure of oxygen and carbon dioxide in the arterial blood, and central chemoreceptors in the brain, which respond to changes in the partial pressure of carbon dioxide in their immediate environment.
Ventilation levels behave as if they were regulated to maintain a constant level of carbon dioxide partial pressure and to ensure adequate oxygen levels in the arterial blood.
Increased activity of chemoreceptors caused by hypoxia or an increase in the partial pressure of carbon dioxide augments both the rate and depth of breathing, which restores partial pressures of oxygen and carbon dioxide to their usual levels.
On the other hand, too much ventilation depresses the partial pressure of carbon dioxide, which leads to a reduction in chemoreceptor activity and a diminution of ventilation.
During sleep and anesthesia, lowering carbon dioxide levels three to four millimeters of mercury below values occurring during wakefulness can cause a total cessation of breathing (apnea).
Peripheral chemoreceptors
Hypoxia, or the reduction of oxygen supply to tissues to below physiological levels (produced, for example, by a trip to high altitudes), stimulates the carotid and aortic bodies, the principal arterial chemoreceptors.
The two carotid bodies are small organs located in the neck at the bifurcation of each of the two common carotid arteries into the internal and external carotid arteries.
This organ is extraordinarily well perfused and responds to changes in the partial pressure of oxygen in the arterial blood flowing through it rather than to the oxygen content of that blood (the amount of oxygen chemically combined with hemoglobin).
The sensory nerve from the carotid body increases its firing rate hyperbolically as the partial pressure of oxygen falls. In addition to responding to hypoxia, the carotid body increases its activity linearly as the partial pressure of carbon dioxide in arterial blood is raised.
This arterial blood parameter rises and falls as air enters and leaves the lungs, and the carotid body senses these fluctuations, responding more to rapid than to slow changes in the partial pressure of carbon dioxide.
Larger oscillations in the partial pressure of carbon dioxide occur with breathing as metabolic rate is increased. The amplitude of these fluctuations, as reflected in the size of carotid body signals, may be used by the brain to detect changes in the metabolic rate and to produce appropriate adjustment in ventilation.
The carotid body communicates with medullary respiratory neurons through sensory fibers that travel with the carotid sinus nerve, a branch of the glossopharyngeal nerve. Microscopically, the carotid body consists of two different types of cells.
The type I cells are arranged in groups and are surrounded by type II cells.
The type II cells are generally not thought to have a direct role in chemoreception. Fine sensory nerve fibers are found in juxtaposition to type I cells, which, unlike type II cells, contain electron-dense vesicles. Acetylcholine, catecholamines, and neuropeptides such as enkephalins, vasoactive intestinal polypeptide, and substance P, are located within the vesicles. It is thought that hypoxia and hypercapnia (excessive carbon dioxide in the blood) cause the release of one or more of these neuroactive substances from the type I cells, which then act on the sensory nerve.
It is possible to interfere independently with the responses of the carotid body to carbon dioxide and oxygen, which suggests that the same mechanisms are not used to sense or transmit changes in oxygen or carbon dioxide.
The aortic bodies located near the arch of the aorta also respond to acute changes in the partial pressure of oxygen, but less well than the carotid body responds to changes in the partial pressure of carbon dioxide. The aortic bodies are responsible for many of the cardiovascular effects of hypoxia.
Central chemoreceptors
Carbon dioxide is one of the most powerful stimulants of breathing. As the partial pressure of carbon dioxide in arterial blood rises, ventilation increases nearly linearly.
Ventilation normally increases by two to four liters per minute with each one millimeter of mercury increase in the partial pressure of carbon dioxide.
Carbon dioxide increases the acidity of the fluid surrounding the cells but also easily passes into cells and thus can make the interior of cells more acidic.
It is not clear whether the receptors respond to the intracellular or extracellular effects of carbon dioxide or acidity.
Even if both the carotid and aortic bodies are removed, inhaling gasses that contain carbon dioxide stimulates breathing. This observation shows that there must be additional receptors that respond to changes in the partial pressure of carbon dioxide.
Current thinking places these receptors near the undersurface (ventral part) of the medulla. However, microscopic examination has not conclusively identified specific chemoreceptor cells in this region.
The same areas of the ventral medulla also contain vasomotor neurons that are concerned with the regulation of blood pressure.
Some investigators suspect that respiratory responses produced at the ventral medullary surface are direct and are caused by interference with excitatory and inhibitory inputs to respiration from these vasomotor neurons.
They further suspect that respiratory chemoreceptors that respond to carbon dioxide are more diffusely distributed in the brain.